Effects of Eutrophication on Fisheries
G. Fred Lee, Ph. D., P.E. and R. Anne Jones, Ph. D.
G. Fred Lee & Associates
El Macero, CA 95618
ABSTRACT: While eutrophication is often discussed in light of its impact on contact recreation, aesthetic character, and the qualitv of a water for water supply, it can also have a marked impact on the fisheries resources of a waterbody. Qualitatively, aquatic plants serve as the foundation for higher trophic level development; aquatic macrophytes provide habitat. Excessive fertility can also, however, have adverse impacts on the quality and quantity of fish that can be supported in a waterbody. This paper presents a discussion of the qualitative impacts of eutrophication and eutrophication management approaches on fisheries. It also introduces an approach for estimating the fish yield that could be sustained in a waterbody, and the change in fish yield that would be expected to result from eutrophication management practices involving P load reduction. This approach, which is an adaptation of the statistical Vollenweider-OECD eutrophication models, is a statistical relationship between the Vollenweider-normalized P loading and wet-weight fish yield. The use of this model as a quantitative management tool in the evaluation of the impact of nutrient load manipulation on the expected yield of fish is discussed.
KEY WORDS: fisheries, eutrophication management, modeling, algae, nutrient control.
I. INTRODUCTION
The overall degree of eutrophication of a waterbody, i.e., the amounts and types of aquatic plants, has a significant impact on the fisheries resources of the waterbody. From a positive point of view, the greater the amount of planktonic algae, typically the greater the sustained fish yield; aquatic macrophytes provide nursery grounds for young fish. On the other hand, there are also negative impacts on the quality of fisheries resources associated with increasing levels of primary production. For a given waterbody, the degree of fertility is, in turn, controlled by the loadings of aquatic plant nutrients to the water-body. This paper reviews the current state of knowledge on the impacts of eutrophication on fisheries resources and the implications of commonly considered eutrophication management practices for fisheries.
It is important to note at the outset that, while the detailed dynamics of any waterbody are complex, the work of Vollenweider,1-3 the authors,4-11 and others has demonstrated that there are some rather simple, fundamental quantitative, stoichiometric relationships between nutrient load and planktonic algal biomass in waterbodies that have universal applicability. These relationships do not purport to describe the details of the ecological interrelationships among trophic levels, nor is that their purpose. Because of their fundamental nature, the extensive data base upon which they were developed, and the extensive documentation of their applicability to a wide variety of types of waterbodies, they are extremely useful for water quality management; they have been demonstrated to provide reliable quantification between nutrient input, which is a characteristic that can be controlled, and planktonic algae-related characteristics. Such a quantification is needed in order to determine how much improvement in water quality or fisheries resources can be obtained for each of the potential management strategies before any are adopted. This paper introduces and focuses on an adaptation of the Vollenweider-OECD eutrophication modeling approach to fisheries resource evaluation and management.
II. CHARACTERISTICS OF EUTROPHICATION
In general terms, eutrophication is the process through which the aquatic plant and other biomass of a waterbody increases. While this process is thus characterized by a continuum of increasing aquatic plant biomass, frequently tne degree of eutrophication that has taken place in a lake or other surface water is described by placing the waterbody in one of two generally qualitative categories: oligotrophic and eutrophic. Although the exact meaning of these two terms is somewhat subjective and depends on the user, it is generally agreed that oligotrophic waterbodies are relatively unproductive and receive comparatively small amounts of aquatic plant nutrients, and eutrophic waterbodies are highly productive in terms of aquatic plants and animals, and experience comparatively high influxes of aquatic plant nutrients.
It is generally found that the more eutrophic a waterbody is, the greater its tendency to experience water quality problems that impair its use as a domestic or industrial water supply, or for contact recreation. In shallow nearshore areas of more eutrophic waterbodies, the aquatic plant growth tends to be manifested as large crops of attached or floating aquatic macrophytes and attached algae. Occasionally, high populations of planktonic algae will also be found in these areas, usually due to wind-induced currents channeling them from the open water to shore. In open waters, aquatic plant growth is usually manifested as planktonic algal growth and in some cases floating aquatic macrophytes. Because of the association of the process of eutrophication with water quality impacts, and because increased aquatic plant growth is associated with increased input of aquatic plant nutrients, the term "eutrophication" is synonymous with "fertilization."
A. Vollenweider-OECD P Load-Eutrophication Response Relationships
During the past decade, in conjunction with the development of management programs for eutrophication-related water quality problems as they affect a variety of beneficial uses of water, considerable progress has been made in the quantification of cause-and-effect relationships that govern eutrophication-related water quality. For most freshwater waterbodies and some nearshore marine waters, phosphorus has been found to be the nutrient that either is limiting aquatic plant growth during periods of water quality concern, or plays a significant role in determining the amount of planktonic algal biomass that the waterbody will support. Its sources are also among the most readily and economically controllable of the aquatic plant nutrients. Thus the loading of phosphorus is generally the most cost-effective focus of nutrient control programs for eutrophication-related water quality management.
Vollenweider1,2 found for a group of water-bodies that the relationship between planktonic algal biomass as measured by chlorophyll and the phosphorus load normalized by waterbody surface area, mean depth, and hydraulic residence time could be expressed as a simple regression. With this foundation and under the leadership of R. Vollenweider and the auspices of the Organization for Economic Corporation and Development (OECD), a multi-nation study was conducted during the mid- to late-1970s on nutrient loading and associated eutrophication response as measured by planktonic algal chlorophyll and related parameters. The relationships between nutrient load and eutrophication response were examined for approximately 200 waterbodies located in North America, Western Europe, Japan, and Australia. Approximately 40 lakes and reservoirs in the U.S. were included in the OECD eutrophication study. Because of the study approach, the load and response data for the U.S. waterbodies were available before those for the rest of the waterbodies evaluated. Following the approach developed by Vollenweider and using the U.S. OECD eutrophication study data base, Rast and Lee10 and Lee et al.8 developed relationships between normalized P load and mean summer chlorophyll, between normalized P load and water clarity as measured by Secchi depth, and between normalized P load and hypolimnetic oxygen depletion rate. The load-response relationships for the approximately 75 OECD waterbodies evaluated in the final OECD eutrophication study report12 published several years later were found to be essentially identical to those that had been developed based on the U.S. OECD eutrophication study data base. The Canadian OECD waterbody data have since been reviewed,13 and the authors and their associates have continued to evaluate the P-load-response relationships for another approximately 40 U.S. waterbodies and about 50 waterbodies in other countries.4,5 They4,5 have found, in accord with the original work of Vollenweider and the U.S. OECD eutrophication study results, statistically significant log-linear regressions between Vollenweider's normalized P loading term and planktonic algal chlorophyll, Secchi depth, and hypolimnetic oxygen depletion rate.
Figure 1 shows the regressions (statistical models) developed by Jones and Lee4 on the basis of data on the U.S. waterbodies evaluated in the U.S. OECD eutrophication study and follow-up studies. The abscissa for these three relationships is Vollenweider's normalized P loading, [(L(P)/qs,)/(l + √t w )], where L(P) is the annual areal P load in mg P/m2/year, t w is the hydraulic residence time (filling time) in years, and qs, is equivalent to the waterbody mean depth divided by the hydraulic residence time, in meters per year. As noted above, Jones and Lee5 expanded the data base for the normalized P loading-chlorophyll relationship by including in the regression the load-response couplings for the additional waterbodies they investigated or as reported in the literature, including the Canadian-OECD waterbodies. Figure 2 shows this updated regression formulated on the basis of more than 325 water-bodies (lakes, reservoirs, and several estuarine systems) around the world and representative of a wide variety of physical, chemical, biological, geographical, and climatological regimes. According to unpublished work of other investigators, another 75 or so waterbodies have been found to follow this relationship as well.
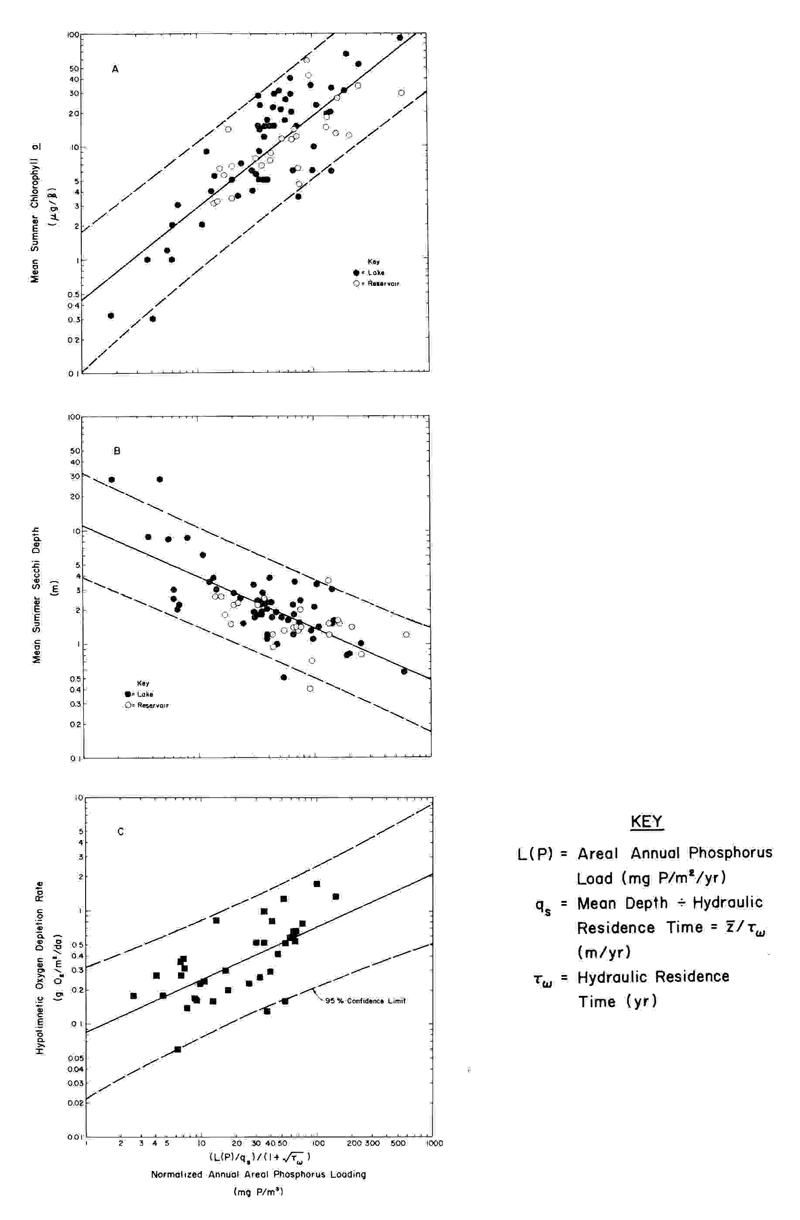
FIGURE 1. Normalized P loading - eutrophication-related water quality response relationships for U.S. quality response relationships for U.S. water bodies - 1982. (After Jones, R.A. and Lee, G.F., J. Water Res. 16, 503, 1982).
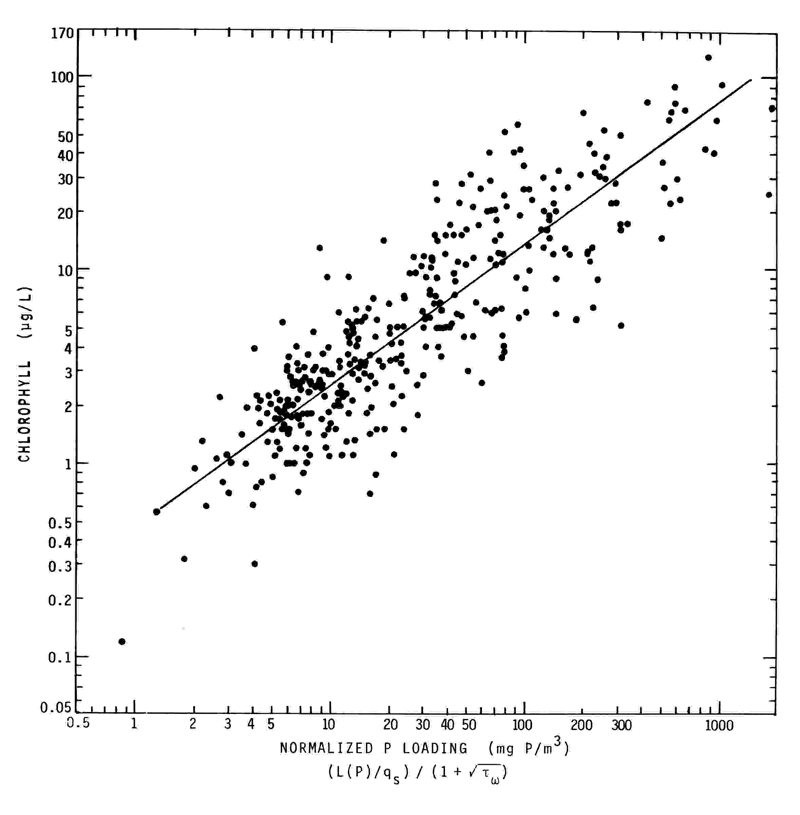
FIGURE 2. Updated Vollenweider-OECD normalized P loading - chlorophyll response relationship. (After Jones, R.A. and Lee, G.F., WHO Water Qual. Bull., 11(2), 67, 1986).
The authors and their associates11 have demonstrated the capability of these models to predict the eutrophication-related water quality characteristics that will result from altering the P load to a waterbody, once the waterbody has approached equilibrium with its new loading. A waterbody is generally considered to have essentially reached equilibrium after a period of time equal to three times the phosphorus residence time following the load change. The P residence time for a waterbody, computed by dividing the mass of P within the waterbody by the annual load of P, is often on the order of 1 year. Thus, the recovery time for many waterbodies is expected to be on the order of 3 years. Also, using the OECD eutrophication study data base and other data, Vollenweider3,12 defined the regression relationship between the normalized P load to lakes and impoundments and planktonic primary production.
While other load-response models have been developed on principles similar to those of the Vollenweider-OECD models, none have the extensive data base or the demonstration of predictive capability that the models described herein have. The use of the Vollenweider-OECD eutrophication modeling approach in conjunction with fisheries resources management is discussed in a subsequent section of this paper.
III. EFFECTS OF EUTROPHICATION ON FISHERIES RESOURCES
Eutrophication of a waterbody can have a number of effects on the quantity and quality of fish within the waterbody. A discussion of major impacts is presented below.
A. Quantity of Fish
One of the most pronounced effects of eutrophication on the fisheries resources of a water-body is the general tendency for an increase in fish stock with an increase in the level of primary production. This was demonstrated by Oglesby,14 who reviewed relationships between fish yield and planktonic algal production. Hanson and Leggett15 reviewed the literature to collect data to examine relationships between fish biomass and yield, and various physical and chemical characteristics of waterbodies. They15 had graphically presented data on about 20 north temperate waterbodies within which they identified a relationship between fish yield and mean epilimnetic total phosphorus concentration. For management purposes, it would be most useful to be able to relate the yield of fish to the principal controllable factor governing algal biomass production, i.e., phosphorus load as normalized by mean depth, hydraulic residence time, and water-body area. Therefore, the authors combined the information provided by Oglesby14 and Hanson and Leggett15 with the relationships shown in Figures 1 and 2 to define the relationship between the Vollenweider-normalized P load to water-bodies and their fish yield.
Oglesby's14 relationship between fish yield and chlorophyll had been developed in part from a relationship between Secchi depth and fish yield and one between Secchi depth and chlorophyll. Since Oglesby's data base for relating Secchi depth to chlorophyll was limited, for those waterbodies for which Secchi depth had been converted to chlorophyll, the authors used the Secchi depth values given by Oglesby and the relationship shown in Figure 1B to estimate the normalized P loads that correspond values, the chlorophyll values and the relationship shown in Figure 2 were used to estimate normalized P loadings. In order to include Hanson and Leggett's data15 in their normalized P loading-fish yield relationship, the authors used the empirical relationship developed by Rast et al.11 between in-lake total P concentration and normalized P loading [(L(P)/qs/(1+√t w )], and thus converted the Hanson and Leggett P concentration values to normalized P loadings.
Since the Hanson and Leggett values for fish yield were for wet weight and since Oglesby had mathematically converted his wet weight values to dry weight for his regressions, the authors chose to represent fish yield as wet weight. While Hanson and Leggett did not indicate the time frame to which the fish yield applied, it was assumed that it was for a 1-year period. After these conversions and appropriate unit adjustments were made, the normalized P loading and fish yield information derived from the data presented by Hanson and Leggett15 were plotted along with those derived from Oglesby14 and are shown in Figure 3. This relationship, which is in keeping with those presented in Figures 1 and 2, forms the foundation of the model that can begin to be used to quantify the effect of altering the P loading to a waterbody on the waterbody's fish yield.
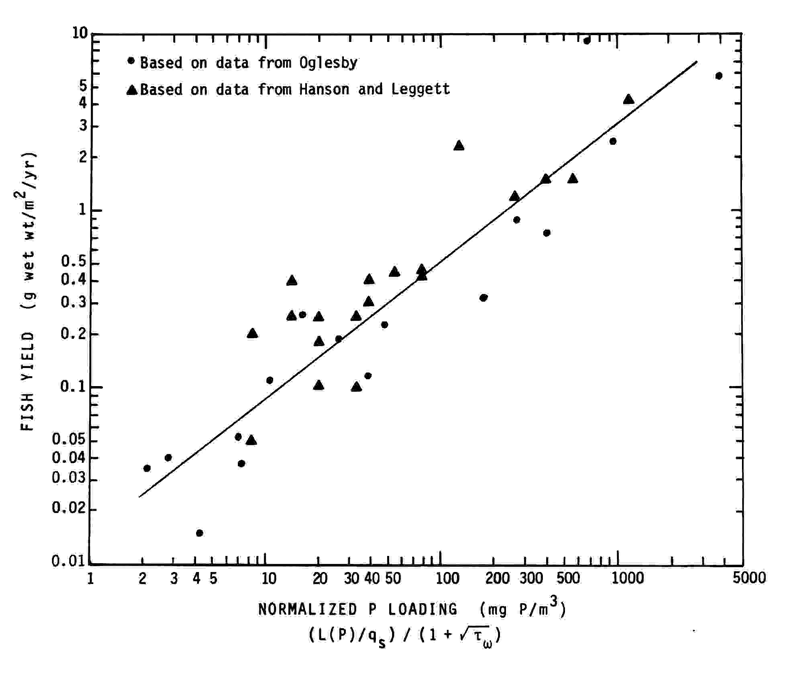
FIGURE 3. Relationship between normalized P loading and fish yield.
Rast et al.11 described in detail how the Vollenweider-OECD models can be used to predict the impact of nutrient load alteration on eutrophication-related water quality characteristics. The same approach may be applied to the model shown in Figure 3. The normalized load-response coordinates for the particular waterbody are plotted on the model; the P load that would result from the load alteration is computed and normalized by the waterbody mean depth, area, and hydraulic residence time according to the Vollenweider-OECD expression; the fish yield that is anticipated to result from the load alteration is determined to be that level corresponding to the intersection in Figure 3 between a vertical line drawn through the new normalized P loading and a line drawn through the original coordinates and parallel to the line of best fit shown in Figure 3. Rast and Lee16 discussed the estimation of nutrient loads for the use of the models for management purposes.
It is now possible, through Vollenweider's original concepts, the results of the OECD and follow-up studies, as well as the work of Oglesby and Hanson and Leggett, to relate quantitatively the Vollenweider-normalized P loading to a waterbody (lake or impoundment) to the water-body's planktonic algal chlorophyll, yearly planktonic primary production, water clarity that is controlled by planktonic algae, hypolimnetic oxygen depletion rate, and fish yield. It is important to emphasize that, despite the myriad factors that affect the dynamics of a waterbody related to fish populations, a relatively simple, fundamental relationship can describe the dependence of overall fish yield on the P loading to the waterbody. The variability about the line of best fit shown in Figure 3 is likely associated with many of these factors. However, because the model is based on the actual relationships between normalized P load and fish yield found for a variety of waterbodies, it also normalizes for many of the factors that may have more subtle impacts on fish yield.
The importance of this relationship is not to disregard the impact of other factors on fish yield. It is rather that a strong, quantitative relationship has been developed between a factor that can be controlled and fish yield, and the fact that such quantification is lacking for most other factors that are qualitatively known to influence fisheries. Furthermore, it gives the water quality manager who is primarily interested in non-fisheries-related aspects of water quality a tool to determine the potential impact of phosphorus load control options on the fisheries resources of a waterbody. Thus, this quantitative tool is of great significance for fisheries management as well as overall water quality management.
As discussed by Rast and Lee,10 Jones and Lee,4 and Rast et al.,11 there are several important factors that must be considered in using these relationships for water quality or fisheries management. First, as with the use of any model, it is important not to blindly plug in values for the morphological and hydrological parameters or the total P load without first examining the shape and characteristics of the waterbody. Jones and Lee4 and Lee and Jones6 discussed those rather unusual situations in which the loading or other factors need to be modified on a site-specific basis in order to apply the models in a technically valid manner. These relationships as currently presented are only applicable to waterbodies in which the P load is primarily manifested as planktonic algae. They would not be applicable to those shallow lakes and reservoirs in which the P load is used primarily to support aquatic macrophytes and attached algae. These relationships also would not be applicable to waterbodies in which there is inordinately high inorganic turbidity or in which allochthonous detritus provides an unusually great proportion of the food source for zooplankton and higher trophic-level organisms. They would not be applicable to waterbodies in which overall fish reproduction is affected by toxic chemicals or lack of adequate habitat, or to some types of "put and take" fisheries. These relationships are, however, applicable to the vast majority of water-bodies and have been demonstrated to be most useful for developing and evaluating management programs.
B. Quality of Fisheries
Eutrophication can also significantly affect the types of fish present in a waterbody. Generally, highly eutrophic waterbodies tend to have dominant populations of rough fish such as carp, which are deemed by most North Americans to be less desirable. While the exact mechanism of the change from more desirable fish to rough fish with increasing fertility is not well understood, it appears to be related to such factors as reduced grazing ability of carnivorous fish brought about by increased turbidity from increased amounts of phytoplankton. This condition tends to be auto-catalytic since once carp and other rough fish become major components of the fish population, they often significantly decrease the overall water clarity through their rooting actions in the water-body sediments.
Some highly eutrophic waterbodies also tend to produce large populations of stunted pan fish. This may be the result of inadequate predation on these fish arising from the inability of predators to see them due to increased turbidity from planktonic algae and suspended sediment. Based on the experience of the authors, stunted pan fish populations tend to occur in waterbodies having average summer Secchi depths of <1 m, which generally correspond to summer average chlorophyll concentrations of 40 to 50 m g/l or more.
Another effect of eutrophication on the quality of fisheries is the elimination of cold water fish from thermally stratified waterbodies that lose the oxygen from their hypolimnia. This oxygen depletion is generally caused by planktonic algae which grow in the surface waters, settle through the thermocline, and become a source of biochemical oxygen demand (BOD) in the hypolimnetic watercolurnn.17 For some waterbodies, ammonia present in the hypolimnion at the onset of thermal stratification exerts a sufficient oxygen demand in nitrification reactions also to reduce significantly the dissolved oxygen concentration in the hypolimnion. Newbry et al.18 found that about one half of the hypolimnetic oxygen depletion in Cherokee Reservoir, part of the Tennessee Valley Authority reservoir system, was due to nitrification; the other half was associated with the bacterial decomposition of phytoplankton.
As mentioned earlier, Rast and Lee10 quantified the regression relationship between nutrient loading and the rate of depletion of oxygen in the hypolimnion of a waterbody that is related to the decomposition of planktonic algae (Figure 1 C). if the characteristics of thermocline development are known so that the dissolved oxygen concentration at the onset of stratification, duration of stratification, and volume of the hypolimnion can be estimated, that relationship can be used to determine the amount of oxygen depletion that would occur in the hypolimnetic waters over the summer, and the lowest dissolved oxygen concentration that would be expected for those conditions. Knowledge of the lowest dissolved oxygen concentration expected will allow an assessment to be made of the potential impact of the planktonic algal biomass supported in the waterbody on the ability of the hypolimnetic waters to sustain fish.
There have been notable cases of significant losses of valuable fisheries apparently due to increases in phosphorus loads to waterbodies that have resulted in increased phytoplankton growths and associated increases in hypolimnetic oxygen depletion rates. The loss of the cold-water cisco population from Lake Mendota, Madison, WI, in the 1930s to 1940s19 is one example of this. Conway20 examined historical information on oxygen depletion rates for Lake Mendota and reported that the hypolimnion of Lake Mendota has gone anoxic every summer since before the turn of the century. Frey19 proposed that, prior to the 1930s, the cisco were surviving the summer hypolimnetic anoxia in a microhabitat just below the thermocline. That area had sufficient oxygen and a suitable temperature to keep the cisco alive during periods of anoxia in the hypolimnion. During that time, the oxygen in that microlayer was maintained by a balance between the rate of 02 diffusion through the thermocline from the epilimnion to the hypolimnion, and hypolimnetic oxygen depletion due to bacterial respiration. In the 1930s and 1940s, there was an increase in the urban and agricultural activities in the lake's watershed that resulted in an increase in the P load to the waterbody. The additional algae produced in response to the increased P load increased the oxygen demand in the hypolimnion sufficiently so that there was no longer enough dissolved oxygen maintained in the microlayer between the epilimnion and hypolimnion to allow the cisco to survive there. Conway20 found that Frey's hypothesis was in accord with the eutrophication history of this lake. As discussed by Conway,20 the loss of cisco from Lake Mendota thus was not a case in which the hypolimnion had remained oxic throughout summers until the 1930s when increased nutrient loads suddenly resulted in hypolimnetic oxygen depletion and elimination of the cisco. Rather, the increased nutrient loadings of the early to mid- 1900s resulted in the elimination of the microhabitat used by the cisco.
It is sometimes alleged that an impact of eutrophication on Lake Erie has been the loss of cold-water fish due to the depletion of oxygen in the hypolimnetic waters. Welch21 reviewed this topic and presented considerable evidence to show that the primary cause of the decline of more desirable, cold-water fish in Lake Erie has been overfishing. A number of studies, such as those of Lee and Jones22 and Vollenweider,23 have shown that, in order to maintain 4 mg/l dissolved oxygen in the hypolimnion of Lake Erie in all years, the P load would not be able to exceed about 1000 to 2000 mt P/year. Since the P load as of the late 1970s was on the order of 20,000 mt P/year, and since the P load could readily and economically have been reduced by only 5000 to 7000 mt P/year, it appears unlikely that cold-water fisheries will ever be maintained in the Central Basin of the lake. Based on the watershed drainage area, the P loads to Lake Erie even prior to the turn of the century were likely to have been sufficient to cause the hypolimnion of Lake Erie to become anoxic in some years, before the onset of the acceleration of the eutrophication of this lake during the past several decades.
The condition of low dissolved oxygen concentrations or anoxia in a waterbody's hypolimnion resulting from the decomposition of plank-tonic algae can also have an adverse effect on the fisheries of downstream waters. As discussed by Krenkel et al.24 and Lee and Jones,17 when masses of anoxic or low-dissolved-oxygen hypolimnetic waters are periodically released from a reservoir, they can eradicate fish populations in the downstream waters. If such waters are released continuously, they can preclude the existence of fish or certain desirable fish in the downstream waters.
Another aspect of eutrophication that can have a significant adverse effect on both the quantity and the quality of fish in a waterbody is winterkill. Winterkill arises principally in shallow waterbodies that develop an ice cover that lasts for several months or more. Under these conditions, respiration of bacteria decomposing the seston, and sediment oxygen demand can cause a decrease in the dissolved oxygen concentration under the ice to levels that could adversely affect fish. This is especially important in shallow lakes because of their limited dissolved oxygen reservoirs. If there is no snow cover over the ice, light can generally penetrate to the water and allow algal growth, and associated with it, the production of sufficient oxygen to maintain fish. However, should there be a snow cover, especially an early snow cover, sufficient to substantially reduce or eliminate light penetration, the dissolved oxygen concentration beneath the ice can be reduced sufficiently to kill fish.
Winterkill tends to be more frequent in eutrophic waterbodies because they support larger amounts of aquatic plant biomass which provides a source of energy for the bacteria. While winterkill can result in complete destruction of all fish in a waterbody, the effect is generally the killing of the more sensitive, often more desirable fish. This situation promotes the dominance of rough fish such as carp which are generally more tolerant of low dissolved oxygen concentrations.
Winterkill problems can generally be eliminated by aeration of the waterbody. It is often possible to add a considerable amount of dissolved oxygen to a waterbody with a relatively small energy input by using a simple diffusion system. Another technique that can be used to add large amounts of oxygen with low energy input is side-stream aeration in which water is pumped from the waterbody to shore where air is added, and then the water is returned to the waterbody. In addition to causing direct absorption of oxygen into the water, these approaches usually make part of the waterbody ice-free and thereby aid in the surface transfer of oxygen from the atmosphere into the water. For those water-bodies prone to winterkill conditions and to developing sufficiently thick coatings of ice to support snow removal equipment, consideration should be given to removing heavy, persistent snow cover so that a substantial part of the ice is essentially snow-free, thereby allowing light penetration through the ice.
If the ordinate in Figure 3 were "Yield of Desirable Fish" rather than the more general "Fish Yield," the relationship for an individual water-body would likely be better represented by a line with two changes in slope. The first decrease in slope would occur at the point corresponding to a sufficient normalized P loading to the particular waterbody to produce sufficient amounts of planktonic algae to result in the reduction of the oxygen concentration in the hypolimnetic waters during the summer to preclude the existence of cold-water species. The second decrease in slope would correspond to the normalized P loading that would be sufficient for the particular waterbody to produce algae in sufficient numbers so as to cause the production of stunted pan fish. Because of waterbody-specific factors that would affect the hypolimnetic oxygen concentration and stunting of fish growth (such as the volume of the hypolimnion for hypolimnetic oxygen depletion), these more finely tuned responses cannot at this time be included in the general representation shown in Figure 3.
It is clear that eutrophication management for optimum fisheries, both in a waterbody and downstream from it, will at times be in conflict with eutrophication management for recreational, domestic and industrial water supply, and other uses of the water since increased nutrient loads lead not only to increased fish production but also to decreased water quality in terms of most non-fisheries-related uses. For optimum cold-water fisheries, there is a balance between having sufficient algal growth for food but not enough to cause detrimental amounts of hypolimnetic oxygen depletion. For any waterbody that thermally stratifies and whose surface waters become too warm for cold-water fish, there is a maximum P load that will provide the maximum yield of cold-water fish. This load may be determined from the information provided in Figures 1C and 3 and the morphological characteristics of the hypolimnion. As discussed by Lee and Jones,22 year to year variations in the position of the thermocline can significantly alter the impact of the P load on the extent of hypolimnetic oxygen depletion.
C. Marine Fisheries
In general, the oceans of the world are nutrient-starved, and their fisheries are limited to a considerable extent by the limited primary production that occurs in them. There are situations, however, in which excessive amounts of nutrients in coastal marine waters can have significant deleterious effects on fisheries and water quality, notably associated with the development of "red tide" organisms, usually dinoflagellates, as well as blooms of other algae that induce oxygen depletion.
The excessive growth of dinoflagellates or phytoplankton in coastal waters can result in the same type of hypolimnetic oxygen depletion discussed previously, associated with bacterial decomposition of the bloom organisms. The coastal waters of the northwest Adriatic Sea in the Emilia-Romagna region of Italy experienced a problem of this type in the mid-1970s when the loadings of nitrogen (apparently primarily derived from domestic, agricultural, and industrial waste) and phosphorus (derived from wastewaters discharged from municipalities located along the coast) resulted in massive phytoplankton blooms in the coastal waters. The oxygen depletion resulting from decomposition of these organisms together with the meteorological conditions that existed during the summer apparently resulted in fish ki1ls.25,26
A similar problem occurred along the New Jersey coast in the summer of 1976 when the waters below the halocline in a large region (approximately 10 to 20 km wide and 150 to 200 km long) became anoxic and caused a massive die-off of benthic fish, shellfish, and other organisms.27 It appears that dinoflagellates that developed in the more open waters of the New York Bight were moved shoreward into a mass as a consequence of the meteorological conditions. These organisms died, settled through the halocline, and their bacterial decomposition depleted the oxygen in the nearshore hypolimnetic waters. The potential exists each year for similar nearshore oxygen depletion to occur.28
During the summer of 1988, there was a massive fish kill in the southern part of Raritan Bay of the Hudson/Raritan Estuary (northern New Jersey coast); an estimated one million fish were killed. According to O'Reilly,29 this fish kill appears to have been caused by the deoxygenation of surface water due to the bacterial decomposition of a massive algal bloom that had occurred in Raritan Bay. Events of this type are expected to occur as long as communities discharging wastewaters to the Hudson/Raritan Estuary do not control phosphorus in their discharges.
Off the coast of Florida, in the coastal waters of Japan, especially the Seto Irdand Sea, as well as in other parts of the world, toxic dinoflagellate blooms have developed, causing fish kills. Japanese fishermen have been particularly impacted by this phenomenon. Millions of kilograms of commercially reared fish (yellowtail) have been lost during single dinoflagellate bloom events. From the information available, it is clear that there is need for work to define the relationship between aquatic plant nutrient input to marine waters and the growth of toxic dinoflagellates which can impact fisheries.
D. Rivers and Streams
As with lakes, impoundments, and estuaries, the more-fertile rivers and streams tend to produce greater fish yields. However, at this time, it is not possible to quantify nutrient load-eutrophication response relationships for flowing waters. While attempts have been made to define these types of relationships, they have had significant technical deficiencies. This is a result of the fact that the loadings of nutrients manifest themselves either as planktonic algae downstream, the location depending on the flow of water and other characteristics of the system, or as attached algae or macrophytes whose biomass is difficult to properly quantify.
One of the primary adverse effects of eutrophication of flowing waters on fisheries is the diel fluctuation in dissolved oxygen concentration associated with photosynthesis and bacterial respiration. In highly fertile streams, early-morning dissolved oxygen concentrations can be sufficiently low for sufficient periods of time to adversely affect fish. This is especially true if there is an additional oxygen demand from allochthonous materials such as from domestic or industrial wastewater sources or upstream reservoir releases. Low dissolved oxygen concentrations can also amplify the effects of certain toxic chemicals such as ammonia that are often discharged into flowing waters. Problems associated with diel fluctuations in dissolved oxygen concentrations can also be found in nearshore waters of lakes, reservoirs, and marine bays.
Eutrophication of flowing waters (or other waters) can also have an impact on fisheries through its impact on the pH of the water. In highly fertile waters, the pH can be raised substantially due to the removal of C02 during photosynthesis; this would occur most frequently in low-alkalinity waters. The impacts that could occur may be direct or indirect. The U.S. EPA water quality criteria and many states' water quality standards stipulate that pH of waters must be maintained between 6.5 and 9. while it is often indicated that pH values above 9 are adverse to fish, there are many excellent eutrophic, warm-water fisheries that routinely have pH values above 9 in the late afternoons during summer months. It is apparent that pHs somewhat above 9 for a period of time do not significantly impair the warm-water fisheries of many waterbodies.
Elevated pH can also have an indirect effect on fisheries due to its effect on the forms of chemicals present. For example, the percent of un-ionized ammonia (the more toxic form of ammonia) increases significantly with increases in pH. This could cause the waters receiving domestic wastewater treatment plant effluent or having another source of ammonia to be in violation of the U.S. EPA's water quality criteria for ammonia published in the July 29, 1985 Federal Register. It is unclear at this time, however, that exceeding the 1-h average or 4-day average criteria values more frequently than once in 3 years as stipulated in the criteria would be of significance to fisheries. This is especially true if these criteria are used as indicated therein for establishing wastewater discharge limitations.
As discussed previously, the eutrophication of an upstream reservoir can have a direct adverse effect on the fisheries potential of a river. Not only can algae be contributed to the stream and contribute to the aforementioned diel dissolved oxygen swing, but also if hypolimnetic waters containing low levels of dissolved oxygen or no dissolved oxygen are released, the types and quantities of fish able to survive in the downstream waters can be significantly adversely affected.
Studies need to be conducted to relate aquatic plant nutrient load to eutrophication response in rivers and streams in order to quantify both beneficial and detrimental effects of increased fertility in lotic systems.
IV. CONTROL OF EXCESSIVE FERTILITY AND FISHERIES IMPLICATIONS
A. Nutrient Load Reduction
Because the input of nutrients, especially P, typically exerts an overwhelming control over the extent of planktonic algal biomass that develops in waterbodies, and because of the readily avail-able and economical approaches for controlling P, one of the most common approaches to eutrophication management is the reduction of the phosphorus load. Lee and Jones9 have reviewed the approaches typically considered for use in North America for nutrient load reduction as part of eutrophication management, as well as their efficacy. Because domestic wastewaters represent a potentially significant source of aquatic plant nutrients to many waterbodies, this source is generally looked to first for control. As discussed by Lee and Jones,9 for a few cents per person served by the treatment plant per day, the phosphorus in domestic wastewaters can typically be reduced on the order of 90%.
The San Joaquin-Sacramento River (Delta) is a water supply source for about 20 million people in California. As discussed by Lee and Jones,34 water utilities that use the Delta as a domestic water supply source frequently have severe algal-caused tastes and odors. Some water utilities are also experiencing increased trihalomethanes (carcinogens) which are correlated with the presence of algae in the water supply reservoir that obtains its water from the Delta. Recently, the authors have determined (Lee and Jones34) that removing phosphorus from the wastewaters that are produced from about 2.5 million people and that are discharged to tributaries of the Delta could significantly reduce planktonic algal growth within the Delta during the summer months. This could, therefore, significantly improve the water quality of the Delta for domestic water supply use. However, there is already considerable concern about the serious declines in some of the fisheries of the Delta. An attempt to limit phosphorus input to the Delta to improve the quality of the domestic water supply would likely be met with opposition from those who are concerned about the potential adverse impacts on fish populations. While, as discussed by Lee and Jones, there can be little doubt that removal of phosphorus from domestic wastewaters added to the Delta will adversely affect the overall fish biomass, especially of rough fish, that develops in the Delta, it is questionable whether such practices would significantly adversely affect the fish populations of greatest concern to the public. The public seems to be primarily concerned with certain species of fish such as striped bass. The changes that apparently have occurred in the populations of these fish are not likely related to fish food limitations. It is possible that the overall trophic state of the Delta could be reduced without significantly adversely affecting the most desirable fish populations in the Delta.
There is an obvious need for better understanding of the factors controlling desirable fish populations in the Delta in order to evaluate whether manipulating phosphorus loads to the Delta could have a significant adverse effect on the fisheries for these fish. If it is found that the significant fish populations within the Delta or that migrate through the Delta are or could be limited by reducing the phosphorus input to the Delta then consideration should be given to assessing the significance of this situation in light of the fact that the current fisheries of the Delta are significantly stimulated by the human population explosion that has occurred in the Delta watershed during the past 50 years. During this period, there has been a very significant increased discharge of phosphorus from domestic waste-water sources to tributaries of the Delta due to the population increase that has occurred in the Delta watershed.
There is a host of non-point sources of nutrients that generally contribute a substantial portion of the total nutrient load to a water-body. These include urban drainage and agricultural and other land runoff. As discussed by Lee and Jones9 and Lee et al.,30 however, substantial portions of nutrients from these sources are unavailable to support algal growth; thus their control would not contribute to the improvement in eutrophication-related water quality. Since non-point-source nutrient control programs are considerably more difficult to develop and implement than most point-source programs, careful evaluation of the availability of non-point-source nutrients should be made and their relative impact on water quality assessed before such measures are implemented.
The overall approach that has generally been used for eutrophication control has been to reduce the loads of phosphorus to the waterbody to the maximum extent possible. This tends to reduce the impairment of recreational use as well as such water supply problems as tastes and odors, increased chlorine demand, shortened filter runs, increased color, and in some situations elevated levels of trihalomethane precursors. However, as noted previously, for a given waterbody, the lower the nutrient load, the lower the planktonic algal production, and the lower the overall fish yield. Thus a conflict exists in making a decision on the degree of eutrophication control desirable for a waterbody for which there is desire to maintain fisheries. Part of eutrophication management for any given waterbody is therefore to determine an "optimum" nutrient load that will provide an appropriate balance of the greatest fish production with minimum significant detrimental eutrophication-related effects, such as loss of cold-water fisheries in the hypolimnion, development of stunted fish populations, and impaired quality of water for domestic or industrial water supply and many recreational uses.
As discussed previously, through the Vollenweider-OECD eutrophication modeling approach, it is now possible to assess quantitatively the impact of given P load reductions on many fisheries-related quality characteristics, such as the oxygen status of hypolimnetic waters, planktonic algae-related water clarity, and overall fish yield, as well as some characteristics important to water supply and recreational uses such as mean and maximum plank-tonic algal chlorophyll concentrations. There are certain more subtle impacts of nutrient loads/eutrophication on fisheries that cannot at this time be reliably predicted, such as the point at which fish become stunted, and loss of fish habitat. For example, it is well known that, as waterbodies become more eutrophic, the depth to which the macrophytes and attached algae develop in nearshore waters tends to decrease. This arises from the increased phytoplankton development reducing light penetration and thereby reducing the light available for the deeper attached and higher plants' development. This results in a loss of fish habitat, especially for young forms. It is possible that decreasing nutrient loads may significantly enhance fisheries for certain types of fish where the development in a waterbody has been limited by aquatic plant growth needed for larval form development. Research is needed to define the relationships between these effects and nutrient load. Once this is done, it will be possible to define better the trade-offs between achieving desirable fisheries-related water quality and desirable water supply and recreational water quality, that are involved in choosing a target nutrient load.
B. Internal Manipulation
There are various in-lake techniques that have been used to reduce the impact of eutrophication on water supply and recreational-use waterbodies. The major control methods of this type and their potential impacts on fisheries-related water quality are discussed below. This discussion is based, in part, on an update of the review by Lee,31 Cooke et al.32 and U.S. EPA33 provide examples of applications of these techniques.
1. Aeration
In attempts to eliminate the problems associated with low-oxygen hypolimnetic water, two forms of in-lake aeration have been practiced. One is complete aeration of the waterbody in which the thermal structure of the waterbody is destroyed. In addition to providing oxygen to the bottom waters, this technique also cools the surface waters. However, it often also results in the significant heating of the hypolimnetic waters so that, depending on the climate of the region, the lake overall could be heated sufficiently so that it becomes impossible to maintain a cold-water fishery. Furthermore, whole-lake aeration mixes the typically nutrient-rich waters of the hypolimnion into the surface waters where they stimulate the growth of more algae during the summer months. Thus, this approach is expected to result in greater levels of algal biomass.
Studies were conducted on Cox Hollow Lake by the Wisconsin Department of Natural Resources to evaluate whether or not whole-lake aeration could increase the fish yield of this waterbody. This lake tended to strongly thermally stratify a few meters below the surface. Since there was complete deoxygenation of the water below the thermocline, the volume of the lake that was available for fish was limited to a relatively narrow band of surface water. Wholelake aeration did not result in a discernible increase in fish yield of this waterbody. The potential value of this approach for improving fisheries or managing eutrophication is still uncertain.
The other approach used for aeration is hypolimnetic aeration in which the thermal structure of the waterbody is maintained. This approach enables the establishment or maintenance of a cold-water fishery in waterbodies whose hypolimnetic oxygen levels become significantly depressed since it provides oxygen to the hypolimnion but does not result in an increase in the hypolimnetic water temperature. Furthermore, since the thermal structure is maintained, this practice does not introduce nutrient-rich hypolimnetic waters into the surface waters. Hypolimnetic aeration with compressed air may result in problems with embolism in fish because it may cause nitrogen supersaturation in the bottom waters. This potential problem can be alleviated by oxygenating with pure oxygen rather than air.
2. Nutrient Removal
One of the more promising techniques for control of excessive fertility in waterbodies whose nutrient inputs cannot be economically controlled to a sufficient extent to effect improvement m water quality is the direct addition of aluminum sulfate (alum) to the waterbody. This technique has been demonstrated in Sweden, Canada (Ontario), the U.S. (Wisconsin), and other locations to effectively remove phosphorus from the water column by sorption of the P onto the aluminum hydroxide floc that forms and then settles to the bottom. This technique also removes algae and color from the water column. Thus far, this technique has proven to significantly reduce algae-related water quality problems without causing a significant direct, observable effect on fish. Of course, any time the primary production in a waterbody is reduced, the overall fish production that the waterbody will sustain over extended periods of time may also be reduced. However, it is possible that, for some waterbodies in which the zooplankters (fish food source) feed primarily on allochthonous detrital material, reduction in the amount of planktonic algae should not affect fish populations. Reduction in planktonic algae, turbidity, and color could result in increased macrophyte growth, perhaps increasing fish nursery grounds. Because this technique does not reduce the input of phosphorus, it has to be practiced periodically depending on the morphology and hydrology of the system.
One of the potential consequences of the addition of alum to a waterbody is the reduction of the waterbody's pH and alkalinity. This would be of greatest concern in low-alkalinity waters such as those influenced by acid rain. One aspect of concern is the potential toxicity of the aluminum added since aluminum is more soluble and hence more available in low-pH waters. It is unlikely that the alum added for P removal for eutrophication control would cause sufficient reduction in pH of alkalinity, or sufficient increase in the available aluminum concentrations to cause pH or toxicity problems. where there is a reason for concern, alkalinity can be raised at the time of treatment by adding lime, or by treating with aluminate instead of alum.
3. Harvesting
Aquatic plant harvesting is a method for removing some of the problems of excessive fertility in some waterbodies; it is widely practiced in several southern states for the control of water hyacinths. In other parts of the country, submerged aquatic macrophytes and attached algae are also effectively harvested by mechanical means. While generally this approach does not significantly alter the nutrient content of the waterbody, it does improve the usability of the water for various recreational purposes, including fishing. Aquatic weed harvesting should be practiced to the minimum extent necessary to permit the use of the waterbody to the extent desired since over-harvesting can lead to the destruction of fish habitat and to the increase in planktonic algal biomass. Since planktonic algae compete with aquatic macrophytes for nutrients, the removal of these macrophytes can induce increases in planktonic algal populations. There have been a number of situations where complete destruction or removal of aquatic macrophytes has resulted in the production of large-scale planktonic algal blooms with their associated water quality problems.
There are several reports in the literature that indicate that attached macrophytes "pump" phosphorus from sediments to the overlying waters. It is possible that in some waterbodies, this pumping action would tend to stimulate plank-tonic algal and floating macrophyte growth to a significant extent beyond that which would occur if the macrophyte populations were substantially reduced. There is need for further work on the role of attached macrophytes in making P available in the water column that would not be available otherwise. Such work could lead to further refinement of the management strategy for aquatic macrophytes in lakes that would optimize fish habitat and minimize eutrophication-related water quality problems.
4. Chemical Treatment
The use of herbicides or other chemicals to control excessive aquatic plant growth can be an effective, although usually an expensive, way to minimize the problems of excessive fertility in waterbodies. This approach, however, can have significant adverse effects on fisheries. Excessive or improper use of chemicals such as copper sulfate can result in toxicity to fish. Also, the widespread use of chemicals of this type throughout a whole waterbody can cause a sufficiently large-scale die-off of aquatic plants to cause the depletion of the oxygen resources of the waterbody as a result of the decay of the vegetation; a fish kill could result. It is imperative that any herbicide be used in a manner so as to minimize or eliminate direct adverse effects on fish such as toxicity, and indirect effects such as oxygen depletion. To accomplish this, the application of herbicides must generally be limited to selected parts of the waterbody and to particular times.
5. Biological Control
One of the potentially important techniques for controlling excessive fertility in waterbodies, but for which the overall water quality implications have not been properly evaluated, is the introduction of plant-eating fish or other animals to graze unwanted macrophytes. Since each type of aquatic plant may have a significantly different impact on water quality, and since fish and other organisms may selectively graze certain types of aquatic plants or zooplankton which, in turn, may selectively graze certain types of plants, the manipulation of fish populations can have a significant impact on the way in which the overall primary production of a waterbody is manifested. It is important to emphasize, however, that it is the aquatic plant nutrients that control the overall fertility. The role of zooplankton and, in turn, fish is to influence how this fertility is manifested. Thus, while the introduction of certain plant-eating fish may reduce the levels of certain macrophytes, the nutrients may then be used for the production of planktonic algae.
State fisheries managers are very aware of the potential impact of the control of rough fish populations on the numbers and types of aquatic plants in waterbodies due to the behavior of the fish as well. Rough fish such as carp stir up the bottom sediments and thereby cause the water to be more turbid which limits the growth of attached algae and macrophytes. With the removal of these fish, the water becomes clearer and these types of aquatic plants can grow. Again, it does not appear, however, that overall production/photosynthesis is drastically changed because of the addition or removal of rough fish. It is generally believed that the effect is a shift from planktonic algal to macrophyte production or vice versa. The reintroduction of carp into a waterbody can readily convert the waterbody from being "weed-choked" back to being relatively weed-free with extensive planktonic algal blooms. A possible exception to this could occur in waterbodies that become sufficiently turbid due to stirring of sediments into the water column so that light limitation precludes limitation of maximum algal biomass production by nutrients. The overall impact of changing from macrophyte to planktonic algal populations, or the reverse, on overall fisheries is not well-known. It can be speculated that the complete elimination of the submerged macrophyte population will greatly reduce the habitat for small fish and eliminate potential breeding areas.
There is also concern about the impact of some of the plant-eating fish used for aquatic macrophyte control, such as grass carp, on other fish. These impacts are of sufficient concern that some states have banned the introduction of these fish into their waters.
There are some who advocate that through the control of fish populations one can significantly alter water quality in a waterbody. The basic premise is that, by removing fish that eat zooplankton, zooplankton are allowed to develop to the maximum extent possible; these zooplankton should then be able to eat much of the phytoplaiikton that have adverse effects on water quality. While intuitively this approach appears to be feasible, its effectiveness on a whole-lake basis remains to be demonstrated. It has no practical applicability, however, to the management of fisheries and it would likely be vigorously opposed by fisheries managers and fishermen.
Another animal that appears to be effective in harvesting macrophytes is the manatee. The use of this animal for this purpose is restricted in the U.S. to the tropical-semitropical marine waters of Florida.
6. Dredging
Dredging has been advocated by some for eutrophication control under the premise that it can remove nutrient-laden sediments that are believed to release nutrients to the overlying waters and stimulate algal growth. However, dredging is not typically useful for reducing internal nutrient load to effect an improvement in eutrophication-related water quality. Nutrient recycling is largely associated with the current year's algae at the sediment surface rather than with sediment-bound P. P released from sediments in thermally stratified waterbodies typically does not affect water quality during critical times of the year because it is trapped beneath the thermocline. Furthermore, dredging is not typically done to sufficient depths to expose nutrient-poor sediments. Dredging will, however, increase the depth of the waterbody. Increasing waterbody depth can decrease the area suitable for attached aquatic macrophyte growth, reduce certain types of fish habitat, and result in increased amounts of planktonic algal growth. The use of dredging should be minimized in waterbodies in which fisheries resources are of concern because of these effects.
Waterbody mean depth is one of the characteristics that affects how a waterbody uses its P load in the production of planktonic algal biomass; it is used to normalize the P loading in the Vollenweider-OECD eutrophication modeling approach. If the overall depth of the waterbody is increased to a sufficient extent, planktonic algal biomass could be decreased to some extent. The impacts of these manipulations are often intricate; in this case, increasing the mean depth could result in an increase in the hydraulic residence time which could negate or diminish the impact of the increased depth on planktonic algal growth. The use of these techniques for altering plank-tonic algal biomass must be evaluated for their potential effectiveness on a site-specific basis using the Vollenweider-OECD models as well as information on fish habitats.
7. Water Level Management
Additional management options that are sometimes available for impoundments are water level management and controlled flushing. Through manipulation of the depth of the water-body and the hydraulic residence time, the planktonic algal chlorophyll production per unit P load can be altered. This is illustrated in the Vollenweider-OECD eutrophication models. As discussed previously, manipulation of water levels can also significandy alter macrophyte and attached algae production.
V. IMPLEMENTATION OF SECTION 314A of PL 92-500
Through the Clean Lakes Program initiated pursuant to Section 3 14A of Public Law 92-500, considerable work has been undertaken to develop control programs for excessively fertile waterbodies. These control programs typically focus on reducing impacts of eutrophication on the use of water for recreation and as an aesthetic asset.
While eutrophication control programs sometimes have some positive impact on fishery resource-related water quality, for example through reducing hypolimnetic oxygen depletion, optimization of fisheries has not been a focal point of 314A activities. As discussed previously, eutrophication control techniques may result in large changes in water quality characteristics as measured by planktonic algal chlorophyll, water clarity, hypolimnetic oxygen depletion rate, macrophyte growth, improved water supply water quality, etc., and at the same time have a limited or detrimental effect on fisheries by removing habitat and nursery areas, and decreasing food supply. The 3 14A activities should in general be expanded to include consideration of the impact of nutrients-eutrophication on fisheries.
The Vollenweider-OECD eutrophication modeling approach provides a technically valid basis for determining the trophic status of a particular waterbody and estimating the water quality benefits that will be derived by controlling the phosphorus load to the waterbody.7 This approach should become one of the bases for judging the appropriateness of 314A activities. The 314A studies should also include collection of information that would allow the further refinement of the normalized P load-fish yield relationship shown in Figure 3, with particular emphasis given to defining the P load to each waterbody that causes a significant shift in the quality of fish.
VI. CONCLUSIONS AND RECOMMENDATIONS
Eutrophication is both beneficial and detrimental to fisheries. Increasing the primary production of a waterbody will generally increase overall fish yield. However, changes in the quality of the fishery to favor those species that are generally less desirable in the North American culture may also be expected to accompany this increase in yield, especially at high trophic levels. One of the most dramatic effects of this type is the loss of cold-water fish associated with deoxygenation of colder, hypolimnetic waters due to bacterial decomposition of algae.
It is recommended that those concerned with eutrophication and fisheries management use the Vollenweider-OECD load-response models as an integral part of evaluating potential management options. They can be used to help develop management approaches for a waterbody to optimize the yield of desirable species but not diminish other aspects of eutrophication-related water quality such as water supply quality, and recreational and aesthetic quality. It is further recommended that research be conducted to quantify the relationship between the degree of fertility of a water-body (normalized nutrient loading) and the quality of fish present using the Vollenweider-OECD modeling approach. Particular attention should be given to the effects of planktonic algal growth on the production of excessive crops of rough fish and stunted fish. As these relationships become better-understood, it should be possible for fisheries managers to refine their recommended management approaches to ensure that the deliberate or de facto fertilization of a waterbody for the purpose of increasing the fish yield is cost-effective and in the best interest of society as a whole in the region of concern.
ACKNOWLEDGMENTS
The original version of this paper was a report prepared at the request of the American Fisheries Society Water Quality Committee. Many individuals, including members of that committee, reviewed drafts of this paper. Their assistance is greatly appreciated. The assistance of Dr. B. Newbry, currently with CH2MHill, Lakewood, CO, in the development of the original version of the normalized P load-fish yield relationship, is also gratefully acknowledged.
REFERENCES
1. Vollenweider, R. A., Scientific fundamentals of the eutrophication of lakes and flowing waters with particular reference to nitrogen and phosphorus as factors in eutrophication, Technical report DASICSII68, OECD, Organization for Economic Cooperation and Development, Paris, 1968.
2. Vollenweider, R. A., Advances in defining critical loading levels for phosphorus in lake eutrophication, Mem. 1st. hal. Idrobiol., 33, 53, 1976.
3. Vollenweider, R. A., Das nahrstoffbelastungs-kon-zept als grundlage fur den externen eingriff in den eutrophierungsprozeB stehender Gewasser und Talsperren, Symp. Wahnbachtalsperrenver-bandes Veroffentlichung der Vortrage, Z. F. Wasser- und Abwasser-Forschung, 12, Jahrgang Nr 2,79, 1979.
4. Jones, R. A. and Lee, G. F., Recent advances in assessing the impact of phosphorus loads on eutrophication-related water quality, J. Water Res., 16, 503, 1982.
5. Jones, R. A. and Lee, G. F., Eutrophication modeling for water quality management: an update of the Vollenweider-OECD model, WHO Water Qual. Bull., 11(2), 67, 1986.
6. Lee, G. F. and Jones, R. A., Evaluation of the effectiveness of phosphorus removal from domestic wastewaters on the eutrophication of receiving waters, presented at Int. Conf. Management Strategies for Phosphorus in the Environment, Lisbon, Portugal, 1985.
7. Lee, G. F. and Jones, R. A., Alternative approach to trophic state classification for water quality management. II. Application of Vollenweider-OECD eutrophication modeling approach, submitted.
8. Lee, G. F., Rast, W., and Jones, R. A., Eutrophication of waterbodies: insights for an age-old problem, Environ. Sci. Technol., 12, 900, 1978.
9. Lee, G. F. and Jones, R. A., The North American experience in eutrophication control, in Proc. Int. Conf. Phosphate, Water and Quality of Life, Paris, France, 1988.
10. Rast, W. and Lee, G. F., Summary analysis of the North American (U.S. portion) OECD eutrophication project: nutrient loading-lake response relationships and trophic state indices, EPA-6O0/3-78-008, U.S. EPA, Corvallis, OR, 1978.
11. Rast, W., Jones, R. A., and Lee, G. F., Predictive capability of U.S. OECD phosphorus loading-eutrophication response models, J. Water Pollui. Control Fed., 55, 990, 1983.
12. Organization for Economic Cooperation and Development (OECD), Eutrophication of waters - monitoring, assessment and control, Final report of the OECD Cooperative Programme on Monitoring of Inland Water (Eutrophication Control), OECD, Paris (97 82 03 1) ISBN 92-64-12298-2, 1982.
13. Janus, L. L. and Vollenweider, R. A., Summary report: the OECD cooperative programme on eutrophication - Canadian contribution, Scientific Series No.131 and 131-S, Canada Centre forlnland Waters, Burlington, Ontario, Canada, 1981.
14. Oglesby, R. T., Relationships of fish yield to lake phytoplankton standing crop, production, and morphoedaphic factors, J. Fish. Res. Board Can., 34, 2271, 1977.
15. Hanson, J. M. and Leggett, W. C., Empirical prediction of fish biomass and yield, Can. J. Fish. Aquat. Sci., 39, 257, 1982.
16. Rast, W. and Lee, G. F., Nutrient loading estimates for lakes, Environ. Eng. Div. ASCE, 109, 502, 1983.
17. Lee, G. F. and Jones, R. A., Mechanisms of the deoxygenation of the hypolimnia of lakes, Occasional Paper, Department of Civil and Environmental Engineering, New Jersey Institute of Technology, Newark, NJ, 1985.
18. Newbry, B. W., Jones, R. A., and Lee, G. F., Assessment and analysis of eutrophication of Tennessee River system impoundments, in Proc. Symp. on Surface Water Impoundments, American Society of Civil Engineers, New York, 1981, 413.
19. Frey, D. G., Distribution ecology of the cisco in Indiana, Invest. Ind. Lakes-Streams, 4, 177, 1955.
20. Conway, C. J., Oxygen Depletion in the Hypolimnion, M.S. thesis, University of Wisconsin-Madison, Madison, 1972.
21. Welch, J., The impact of inorganic phosphates in the environment, U.S. EPA Office of Toxic Substances, EPA 560/1-78-003, Washington, D.C., 1978.
22. Lee, G. F. and Jones, R. A., Water quality characteristics of the US waters of Lake Ontario during the IFYGL, and modeling contaminant load-water quality response relationships in the nearshore waters of the Great Lakes, Contract report (Project No.03-~22-35196) submitted to NOAA, Ann Arbor, MI, 1979.
23. Vollenweider, R. A., Canada Centre for Inland Waters, Burlington, Ontario, Canada, personal communication to G. Fred Lee, 1979.
24. Krenkel, P. A., Lee, G. F., and Jones, R. A., Effects of TVA impoundments on downstream water quality and biota, in The Ecology of Regulated Streams, Ward, J. V. and Stanford, J. A., Eds., Plenum Press, New York, 1979, 289.
25. Lee, G. F. and Jones, R. A., An approach for the development of a eutrophication control program for the Emilia-Romagna coastal waters of the Adriatic Sea, Occasional Paper No.27, Department of Civil and Environmental Engineering, New Jersey Institute of Technology, Newark, NJ, 1977.
26. Lee, G. F., Indicazioni per lo sviluppo di un programma di controllo dell'eutrofizzazione delle acque costiere adriatiche della regione Emrria-Romagna, Proc. Emilia-Romagna Conf. on Eutrophication of Northwest Nearshore Adriatic Sea, 1978, 100.
27. Swanson, R. L. and Sindermann, C. J., Eds., Oxygen Depletion and Associated Benthic Mortalities in New York Bight, 1976, NOAA Professional Paper 11, National Oceanic and Atmospheric Administration, U.S. Government Printing Office, Washington, D.C., 1979.
28. Lee, G. F. and Jones, R. A., Impact of reducing P loads from domestic wastewaters on algal-related water quality of northern New Jersey's coastal waters, Report to New Jersey Marine Sciences Consortium, Fort Hancock, NJ, 1987.
29. O'Reilly, J., National Marine Fisheries Service Sandy Rook Laboratory, Fort Hancock, NJ, personal communication to G. Fred Lee, 1988.
30. Lee, G. F., Jones, R. A., and Rast, W., Availability of phosphorus to phytoplankton and its implications for phosphorus management strategies, in Phosphorus Management Strategies for Lakes, Ann Arbor Press, Ann Arbor, MI, 1980, 259.
31. Lee, G. F., Eutrophication, Trans. Northeast Fish Wildl. Conf., p.39, 1973.
32. Cooke, G. D., Welch, E. B., Peterson, Sp. A., and Newroth, P. R., Lake and Reservoir Restoration, Butterworths, Boston, 1986.
33. U.S. EPA, Restoration of lakes and inland waters, Proc. Int. Symp. on Inland Waters and Lake Restoration, EPA 440/5-81-010, U.S. EPA Office of Water Regulation and Standards, Washington, D.C., December 1980.
34. Lee, G. F. and Jones, R. A., Managing Delta algal related drinking water quality: tastes and odors and THM precursors, Proc. Conf. Protecting Water Supply Water Quality at the Source, University of California Water Resources Center, Davis, CA, in press.
Reference as: "Lee, G. F. and Jones, R. A., 'Effects of Eutrophication on Fisheries,' Reviews in Aquatic Sciences, 5:287-305, CRC Press, Boca Raton, FL (1991)."

|
|